Устьица у растения: определение, расположение, функции. Значение устьиц в дыхании растений
Устьица представляют собой высокоспециализированные образования эпидермы, состоящие из двух замыкающих клеток, между которыми имеется своеобразный межклетник, или устьичная щель ( рис. 37). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Под щелью располагается дыхательная, или воздушная, полость, окруженная клетками мякоти листа . Клетки эпидермы, примыкающие к замыкающим, получили название побочных, или околоустьичных. Они участвуют в движении замыкающих клеток. Замыкающие и побочные клетки образуют устьичный аппарат. Число побочных клеток и их расположение относительно устьичной щели позволяют выделять ряд устьичных типов. Их изучением занимается стоматография. Данные стоматографии нередко используются в систематике растений для уточнения систематического положения таксонов. Наиболее часто встречающиеся устьичные типы показаны на рисунке 38 .
Аномоцитный тип устьичного аппарата обычен для всех групп высших растений , исключая хвойные . Побочные клетки в этом случае не отличаются от остальных клеток эпидермы. Диацитный тип характеризуется только двумя побочными клетками, общая стенка которых перпендикулярна устьичной щели. Этот тип обнаружен у некоторых цветковых , в частности у большинства губоцветных и гвоздичных . При парацитном типе побочные клетки располагаются параллельно замыкающим и устьичной щели. Он найден у папоротников , хвощей и ряда цветковых растений . Анизоцитный тип обнаружен только у цветковых растений . Здесь замыкающие клетки окружены тремя побочными, одна из которых заметно крупнее или мельче остальных. Тетрацитным типом устьичного аппарата характеризуются преимущественно однодольные . При энциклоцитном типе побочные клетки образуют узкое кольцо вокруг замыкающих клеток. Подобная структура найдена у папоротников , голосеменных и ряда цветковых . Расположение замыкающих клеток относительно прочих клеток эпидермы у разных видов неодинаково. В одних случаях замыкающие клетки находятся на одном уровне с эпидермальными, иногда выступают над ними или, напротив, залегают значительно глубже (погруженные устьица). Последнее наблюдается у растений, приспособленных к засушливым условиям. Иногда углубления, в которых располагаются устьица, выстланы или прикрыты волосками. Называются они устьичными криптами .
Число и распределение устьиц на листе или побеге варьируют в зависимости от вида растений и условий жизни. Число их обычно колеблется от нескольких десятков до нескольких сотен на 1 кв.мм поверхности.
Механизм движения замыкающих клеток весьма сложен и неодинаков у разных видов. У большинства растений при недостаточном водоснабжении в ночные часы, а иногда и днем тургор в замыкающих клетках понижается и щель замыкается, снижая тем самым уровень транспирации. С повышением тургора устьица открываются. Считают, что главная роль в этих изменениях принадлежит ионам калия. Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов . Первичный крахмал хлоропластов , превращаясь в сахар , повышает концентрацию клеточного сока. Это способствует притоку воды из соседних клеток и переходу замыкающих клеток в упругое состояние.
Общая площадь устьичных отверстий составляет лишь 1-2% площади листа. Несмотря на это, транспирация при открытых устьичных щелях достигает 50-70% испарения, равного по площади открытой водной поверхности.
Клетки эпидермиса почти непроницаемы для воды и газов благодаря своеоб-разному устройству их наружной стенки. Как же осуществляются газообмен между растением и наружной средой и испарение воды — процессы, не-обходимые для нормальной жизнедеятельности растения? Среди клеток эпидермиса встречаются характерные образования, называемые устьи-цами .
Устьице — щелевидное отверстие, окаймленное с двух сторон двумя замыкающими клетками, имеющими большей частью полулун-ную форму.
Клетки эти живые и со-держат хлорофилловые зерна и кру-пинки крахмала, отсутствующие в других клетках эпидермиса. Особен-но много устьиц на листе. Вид усть-ица с поверхности и в разрезе изоб-ражен на рисунке 40. На поперечном разрезе видно, что непосредственно под устьицем внутри ткани листа находится полость, называемая дыхательной . В пределах щели за-мыкающие клетки более сближены в средней части клеток, а выше и ниже они дальше отступают друг от друга, образуя пространства, называемые передним и задним дво-риком .
Замыкающие клетки способны увеличивать и сокращать свои разме-ры, благодаря чему устьичная щель то широко раскрывается, то сужива-ется или даже совсем бывает закрыта.
Таким образом, замыкающие клетки являются аппаратом, регулирующим процесс открывания и закрывания устьиц.
Как же осуществляется этот процесс?
Стенки замыкающих клеток, обращенные к щели, утолщены значи-тельно сильнее, чем стенки, обращенные к соседним клеткам эпидер-миса. Когда растение освещено и имеет избыток влаги, в хлорофилловых зернах замыкающих клеток происходит накопление крахмала, часть ко-торого превращается в сахар. Сахар, растворенный в клеточном соке, при-тягивает воду из соседних клеток эпидермиса, вследствие чего в замы-кающих клетках повышается тургор. Сильное давление приводит к выпячиванию стенок клеток, примыкающих к эпидермальным, а про-тивоположные, сильно утолщенные стенки выпрямляются. Вследствие этого устьичная щель раскрывается, и газообмен, а также испарение воды увеличиваются. В темноте или при недостатке влаги тур-горное давление уменьшается, замыкающие клетки принимают прежнее по-ложение и утолщенные стенки смыкаются. Щель устьица закрывается.
Устьица расположены на всех молодых неодревесневших наземных ор-ганах растения. Особенно много их на листьях, причем здесь они располо-жены, главным образом, на нижней поверхности. Если лист расположен вертикально, то устьица развиваются с обеих его сторон. У плавающих на поверхности воды листьев некоторых водных растений (например, кувшинки, кубышки) устьица расположены только на верхней стороне листа. Материал с сайта
Число устьиц на 1 кв. мм листовой поверхности в среднем равно 300, однако иногда достигает 600 и более. У рогоза (Typha ) насчитывают свыше 1300 устьиц на 1 кв. мм. Листья, погруженные в воду, устьиц не имеют. Расположены устьица чаще всего равномерно по всей поверхности кожи-цы, но у некоторых растений собраны группами. У однодольных растений, а также на хвоинках многих хвойных они расположены продольными рядами. У растений засушливых областей неред-ко устьица бывают погружены в ткань листа.
Развитие устьиц обычно происхо-дит следующим образом. В отдельных клетках эпидермиса образуются дуго-образные стенки, разделяющие клетку на несколько более мелких так, что центральная из них становится родо-начальницей устьиц. Эта клетка раз-деляется продольной (по оси клетки) перегородкой. Затем эта перегородка расщепляется, и образуется щель. Ог-раничивающие ее клетки становятся замыкающими клетками устьица. У не-которых печеночных мхов имеются свое-образные устьица, лишенные замыкаю-щих клеток.
На этой странице материал по темам:
Лист - это вегетативный орган растений, является частью побега. Функции листа - фотосинтез, испарение воды (транспирация) и газообмен. Кроме этих основных функций, в результате идиоадаптаций к различным условиям существования листья, видоизменяясь, могут служить следующим целям.
- Накопления питательных веществ (лук, капуста), воды (алоэ);
- защиты от поедания животными (колючки кактуса и барбариса);
- вегетативного размножения (бегония, фиалка);
- улавливания и переваривания насекомых (росянка, венерина мухоловка);
- движения и укрепления слабого стебля (усики гороха, вики);
- удаления продуктов обмена веществ во время листопада (у деревьев и кустарников).
Общая характеристика листа растения
Листья у большинства растений зеленые, чаще всего - плоские, обычно двустороннесимметричные. Размеры от нескольких миллиметров (ряска) до 10-15м (у пальм).
Лист формируется из клеток образовательной ткани конуса нарастания стебля. Зачаток листа дифференцируется на:
- Листовую пластинку;
- черешок, с помощью которого лист прикрепляется к стеблю;
- прилистники.
У некоторых растений черешков нет, такие листья в отличие от черешковых называются сидячими . Прилистники также бывают не у всех растений. Они представляют собой различных размеров парные придатки у основания черешка листа. Форма их разнообразна (пленки, чешуйки, маленькие листочки, колючки), функция - защитная.
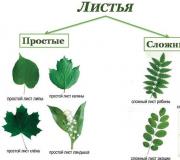
Простые и сложные листья различают по числу листовых пластинок. Простой лист имеет одну пластинку и отпадает целиком. У сложного на черешке располагается несколько пластинок. Они прикрепляются к главному черешку своими маленькими черешочками и называются листочками. При отмирании сложного листа сначала отпадают листочки, а затем - главный черешок.

Листовые пластинки разнообразны по форме: линейные (злаки), овальные (акации), ланцетовидные (ива), яйцевидные (груша), стреловидные (стрелолист) и т.д.
Листовые пластинки в разных направлениях пронизаны жилками, которые представляют собой сосудисто-волокнистые пучки и придают листу прочность. У листьев двудольных растений чаще всего сетчатое или перистое жилкование, а у листьев однодольных - параллельное или дуговое.
Края листовой пластинки могут быть сплошными, такой лист называется цельнокрайним (сирень) или с выемками. В зависимости от формы выемки, по краю листовой пластинки различают листья зубчатые, пильчатые, городчатые и др. У зубчатых листьев зубцы имеют более или менее равные стороны (бук, лещина), у пильчатых - одна сторона зубца длиннее другой (груша), городчатые - имеют острые выемки и тупые выпуклости (шалфей, будра). Все эти листья называются цельными, так как выемки у них неглубокие, не достигают ширины пластинки.

При наличии более глубоких выемок листья бывают лопастные, когда глубина выемки равна половине ширины пластинки (дуб), раздельные - более половины (мак). У рассеченных листьев выемки доходят до средней жилки или до основания листа (репейник).
В оптимальных условиях роста нижние и верхние листья побегов неодинаковы. Различают низовые, срединные и верховые листья. Такая дифференцировка определяется еще в почке.
Низовые, или первые, листья побега - это чешуйки почек, наружные сухие чешуи луковиц, семядольные листья. Низовые листья при развитии побега обычно опадают. К низовым относят и листья прикорневых розеток. Срединные, или стебельные, листья типичны для растений всех видов. Верховые листья обычно имеют более мелкие размеры, располагаются вблизи цветков или соцветий, бывают окрашены в различные цвета, либо бесцветны (кроющие листья цветков, соцветий, прицветники) .
Типы расположения листов
Существует три основных типа листорасположения:
- Очередное или спиральное;
- супротивное;
- мутовчатое.
При очередном расположении одиночные листья прикрепляются к стеблевым узлам по спирали (яблоня, фикус). При супротивном - два листа в узле располагаются один против другого (сирень, клен). Мутовчатое листорасположение - три и более листа в узле охватывают стебель кольцом (элодея, олеандр).
Любое листорасположение позволяет растениям улавливать максимальное количество света, так как листья образуют листовую мозаику и не затеняют друг друга.

Клеточное строение листа
Лист, как и все другие органы растения, имеет клеточное строение. Верхняя и нижняя поверхности листовой пластинки покрыты кожицей. Живые бесцветные клетки кожицы содержат цитоплазму и ядро, располагаются одним сплошным слоем. Наружные оболочки их утолщены.
Устьица — органы дыхания растения
В кожице находятся устьица - щели, образованные двумя замыкающими, или устьичными, клетками. Замыкающие клетки имеют полулунную форму и содержат цитоплазму, ядро, хлоропласты и центральную вакуоль. Оболочки этих клеток утолщены неравномерно: внутренняя, обращенная к щели, толще, чем противоположная.

Изменение тургора замыкающих клеток меняет их форму, благодаря чему устьичная щель бывает открыта, сужена или полностью закрыта в зависимости от условий окружающей среды. Так, днем устьица открыты, а ночью и в жаркую сухую погоду - закрыты. Роль устьиц заключается в регуляции испарения воды растением и газообмена с окружающей средой.
Устьица располагаются обычно на нижней поверхности листа, но бывают и на верхней, иногда они распределены более или менее равномерно по обе стороны (кукуруза); у водных плавающих растений устьица расположены только на верхней стороне листа. Число устьиц на единице площади листа зависит от вида растений, условий роста. В среднем их 100-300 на 1мм 2 поверхности, но может быть и значительно больше.
Мякоть листа (мезофил)
Между верхней и нижней кожицей листовой пластинки располагается мякоть листа (мезофил). Под верхним слоем находится один или несколько слоев крупных прямоугольных клеток, которые имеют многочисленные хлоропласты. Это столбчатая, или палисадная, паренхима - основная ассимиляционная ткань, в которой осуществляются процессы фотосинтеза.
Под палисадной паренхимой находится несколько слоев клеток неправильной формы с большими межклетниками. Эти слои клеток образуют губчатую, или рыхлую, паренхиму. В клетках губчатой паренхимы содержится меньше хлоропластов. Они выполняют функции транспирации, газообмена и запасания питательных веществ.
Мякоть листа пронизана густой сетью жилок, сосудисто-волокнистых пучков, осуществляющих снабжение листа водой и растворенными в ней веществами, а также отведение из листа ассимилянтов. Кроме того, жилки выполняют механическую роль. По мере отхода жилок от основания листа и приближения их к вершине, они утончаются за счет ветвления и постепенного выпадения механических элементов, затем ситовидных трубок, наконец, трахеид. Мельчайшие разветвления у самого края листа обычно состоят только из трахеид.
 Схема строения листа растения
Схема строения листа растения
Микроскопическое строение листовой пластинки существенно меняется даже в рамках одной систематической группы растений, в зависимости от разных условий произрастания, прежде всего, от условий освещения и водоснабжения. У растений затененных мест часто отсутствует палисадная перенхима. Клетки ассимиляционной ткани имеют более крупные палисады, концентрация хлорофилла в них выше, чем у светолюбивых растений.
Фотосинтез
В хлоропластах клеток мякоти (особенно столбчатой паренхимы) на свету происходит процесс фотосинтеза. Сущность его заключается в том, что зеленые растения поглощают солнечную энергию и из углекислого газа и воды создают сложные органические вещества. В атмосферу при этом выделяется свободный кислород.
Созданные зелеными растениями органические вещества являются пищей не только для самих растений, но и для животных и человека. Таким образом, жизнь на земле зависит от зеленых растений.
Весь кислород, содержащийся в атмосфере, имеет фотосинтетическое происхождение, он накапливается за счет жизнедеятельности зеленых растений и его количественное содержание благодаря фотосинтезу поддерживается постоянным (около 21%).
Используя углекислый газ из атмосферы для процесса фотосинтеза, зеленые растения тем самым очищают воздух.
Испарение воды листьями (транспирация)
Кроме фотосинтеза и газообмена в листьях происходит процесс транспирации - испарения воды листьями. Основную роль в испарении выполняют устьица, частично в этом процессе принимает участие и вся поверхность листа. В связи с этим различают устьичную транспирацию и кутикулярную - через поверхность кутикулы, покрывающей эпидермис листа. Кутикулярная транспирация значительно меньше устьичной: у старых листьев 5-10% общей транспирации, однако у молодых листьев, имеющих тонкую кутикулу, может достигать 40-70%.
Поскольку транспирация осуществляется в основном через устьица, куда проникает и углекислый газ для процесса фотосинтеза, существует взаимосвязь между испарением воды и накоплением сухого вещества в растении. Количество воды, которое испаряется растением для построения 1г сухого вещества, называется транспирационным коэффициентом . Величина его колеблется от 30 до 1000 и зависит от условий роста, вида и сорта растений.
На построение своего тела растение использует в среднем 0,2% пропускаемой воды, остальная расходуется на терморегуляцию и транспорт минеральных веществ.
Транспирация создает сосущую силу в клетке листа и корня, поддерживая тем самым постоянное передвижение воды по растению. В связи с этим листья получили название верхнего водяного насоса в отличие от корневой системы - нижнего водяного насоса, который нагнетает воду в растение.
Испарение защищает листья от перегревания, что имеет большое значение для всех процессов жизнедеятельности растения, особенно - фотосинтеза.
Растения засушливых мест, а также в сухую погоду испаряют больше воды, чем в условиях повышенной влажности. Регулируется испарение воды кроме устьиц защитными образованиями на кожице листа. Эти образования: кутикула, восковой налет, опушение из различных волосков и др. У растений-суккулентов лист превращается в колючки (кактусы), а его функции выполняет стебель. Растения влажных мест обитания имеют крупные листовые пластинки, на кожице нет защитных образований.
При затрудненном испарении у растений наблюдается гуттация - выделение воды через устьица в капельно-жидком состоянии. Это явление происходит в природе обычно утром, когда воздух приближается к насыщению водяными парами, или перед дождем. В условиях лаборатории гуттацию можно наблюдать, накрыв молодые проростки пшеницы стеклянными колпаками. Через короткий срок на кончиках их листьев появляются капельки жидкости.
Система выделения — опадание листьев (листопад)
Биологическим приспособлением растений к защите от испарения является листопад - массовое опадение листьев на холодное или жаркое время года. В умеренных зонах деревья сбрасывают листья на зиму, когда корни не могут подавать воду из замерзшей почвы, а мороз иссушает растение. В тропиках листопад наблюдают в сухой период года.

Подготовка к сбрасыванию листьев начинается при ослаблении интенсивности жизненных процессов в конце лета - начале осени. Прежде всего происходит разрушение хлорофилла, другие пигменты (каротин и ксантофилл) сохраняются дольше и придают листьям осеннюю окраску. Затем у основания черешка листа паренхимные клетки начинают делиться и образуют отделительный слой. После этого лист отрывается, а на стебле остается след - листовой рубец. Ко времени листопада листья стареют, в них скапливаются ненужные продукты обмена веществ, которые удаляются из растения вместе с опавшими листьями.
Все растения (обычно это деревья и кустарники, реже - травы) делятся на листопадные и вечнозеленые. У листопадных листья развиваются в течение одного вегетационного сезона. Ежегодно с наступлением неблагоприятных условий они опадают. Листья вечнозеленых растений живут от 1 до 15 лет. Отмирание части старых и появление новых листьев происходит постоянно, дерево кажется вечнозеленым (хвойные, цитрусовые).
И выделяет углекислоту . Кроме того, своими зелеными частями оно поглощает углекислоту и выделяет кислород. Затем, растение постоянно испаряет воду. Так как кутикула , которой покрыты листья и молодые стебли, очень слабо пропускает через себя газы и водяные пары, то для беспрепятственного обмена с окружающей атмосферой в кожице имеются особые отверстия, называемые У. На поперечном разрезе листа (фиг. 1) У. представляется в виде щели (S ), ведущей в воздухоносную полость (i ).
Фиг. 1. Устьице (S ) листа гиацинта в разрезе.
По обеим сторонам У. находится по одной замыкающей клетке. Оболочки замыкающих клеток дают в сторону устьичного отверстия по два выроста, благодаря которым оно распадается на две камеры: передний и задний дворик. При рассматривании с поверхности - У. представляется в виде продолговатой щели, окруженной двумя полулунными замыкающими клетками (фиг. 2).
Фиг. 2. Устьице листа Sedum purpurascens с поверхности.
Днем У. открыты, на ночь же они закрываются. У. закрываются также днем во время засухи. Закрывание У. производится замыкающими клетками. Если кусочек кожицы листа положить в воду, то У. продолжают оставаться открытыми. Если же воду заменить раствором сахара, вызывающим плазмолиз клеток, то У. закроются. Так как плазмолиз клеток сопровождается уменьшением их объема, то отсюда следует, что закрывание У. есть результат уменьшения объема замыкающих клеток. Во время засухи замыкающие клетки теряют часть своей воды, уменьшаются в объеме и закрывают У. Лист оказывается покрытым сплошным слоем кутикулы , слабо пропускающей водяные пары, чем и предохраняется от дальнейшего высыхания. Ночное закрывание У. объясняется следующими соображениями. Замыкающие клетки постоянно содержат в себе хлорофилловые зерна и поэтому способны к усвоению атмосферной углекислоты, т. е. к самостоятельному питанию. Накопленные на свету органические вещества сильно притягивают к себе из окружающих клеток воду, поэтому замыкающие клетки увеличиваются в объеме и открываются. Ночью же выработанные на свету органические вещества расходуются, а вместе с ними утрачивается способность притягивать воду, и У. закрываются. У. находятся как на листьях, так и на стеблях. На листьях они помещаются или на обеих поверхностях, или же на одной из них. Травянистые, мягкие листья имеют У. как на верхней, так и на нижней поверхности. Твердые кожистые листья имеют У. почти исключительно на нижней поверхности. У листьев, плавающих на поверхности воды, У. исключительно находятся на верхней стороне. Количество У. у различных растений очень различно. Для большинства листьев число У., находящихся на одном квадратном миллиметре, колеблется между 40 и 300. Наибольшее число У. находится на нижней поверхности листа Brassica Rара - на 1 кв. мм 716. Существует некоторая зависимость между количеством У. и влажностью места. В общем растения влажных местностей имеют более У., чем растения сухих местностей. Кроме обыкновенных У., служащих для газового обмена, у многих растений имеются еще водяные У. Они служат для выделения воды не в газообразном состоянии, но в жидком. Вместо лежащей под обыкновенными У. воздухоносной полости под водяными У. находится особая водоносная ткань, состоящая из клеток с тонкими оболочками. Водяные У. встречаются по большей части у растений сырых местностей и находятся на различных частях листьев независимо от находящихся тут же обыкновенных У. Водяные У. выделяют капли воды по большей части тогда, когда вследствие большой влажности воздуха не могут испарять воду воздухоносные У. Кроме водяных У., существует целый ряд разнообразных приспособлений для выделения листьями воды в жидком виде. Все такие образования получили название гидатод (Hydathode). Примером могут служить гидатоды Gonocaryum pyriforme (фиг. 3).
Фиг. 3. Гидатода листа Gonocaryum pyriforme.
На поперечном разрезе через лист видно, что некоторые из клеток кожицы особенным образом изменились и превратились в гидатоды. Каждая гидатода состоит из трех частей. Наружу выдается идущий вкось вырост, пронизанный узким канальцем, через который вытекает вода гидатоды. Средняя часть имеет вид воронки с очень утолщенными стенками. Нижняя часть гидатоды состоит из тонкостенного пузыря. Некоторые растения выделяют своими листьями большие количества воды, не имея никаких особо устроенных гидатод. Напр. различные виды Salacia выделяют между 6-7 часами утра такие большие количества воды, что вполне заслуживают названия дождевых кустарников: при легком прикосновении к ветвям с них падает настоящий дождь. Вода выделяется простыми порами, покрывающими в большом количестве наружные оболочки клеток кожицы.
Для соприкосновения листа с атмосферой имеются поры - устьица. Устьице - это отверстие (щель), ограниченная двумя замыкающими клетками. Устьица встречаются у всех наземных органов растения, но больше всего у листьев. Каждая замыкающая клетка устьица в отличие от клеток эпидермиса имеет хлоропласта. В них происходит фотосинтез, хотя с меньшей интенсивностью, чем в клетках мезофилла. Устьица - одно из оригинальных приспособлений, обладающих способностью открываться и закрываться в зависимости от насыщенности замыкающих клеток водой . Обычно устьичные отверстия ограничены двумя замыкающими клетками , стенки которых неравномерно утолщены. У двудольных растений замыкающие клетки бобовидной, или полулунной, формы, при этом их внутренние прилегающие друг к другу клеточные стенки более толстые, а внешние - более тонкие. Протопласты замыкающих клеток связаны в единое целое перфорациями в основании граничащих общих стенок. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на клеточные стенки, и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель.
В последнее время доказано, что для движения устьиц большое значение имеет также расположение микрофибрилл целлюлозы . Если обычно в клетках листьев целлюлозные фибриллы ориентированы в длину и в этом направлении утолщены, то в замыкающих клетках устьиц микрофибриллы организованы радиально, что усиливает устойчивость к процессу растяжения.
У злаков строение замыкающих клеток несколько иное. Они представлены двумя удлиненными клетками, на концах которых стенки более тонкие. При насыщении водой более тонкие стенки на концах растягиваются и раздвигают замыкающие клетки, благодаря чему образуется щель.
Число устьичных отверстий колеблется в зависимости от вида растений от 10 до 600 на 1 мм2 листа. У многих растений (75% видов), в том числе для большинства древесных, устьица расположены на нижней стороне листа. Диаметр устьичных щелей составляет всего 3-12 мкм. Устьица соединяют внутренние пространства листа с внешней средой. Вода поступает в лист через сеть жилок, в которых расположены сосудистые элементы. Возможны три пути испарения:
- через устьица - устьичная,
- кутикулу - кутикулярная,
- через чечевички - лентикулярная транспирация.
Впервые разграничение на кутикулярную и устьичную транспирацию было введено в 1877 г.
Основные типы устьичного аппарата листа растений.
- аномоцитный (у всех высших растений, кроме хвощей),
- диацитный (у папоротникови цветковых),
- парацитный (у папоротников, хвощей, цветковых и гнетовых),
- анизоцитный (только у цветковых),
- тетрацитный (главным образом у однодольных),
- энциклоцитный (у папоротников, голосеменныхи цветковых).
Устьица представляют собой высокоспециализированные образования эпидермы, состоящие из двух замыкающих клеток, между которыми имеется своеобразный межклетник, или устьичная щель.
Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Под щелью располагается дыхательная, или воздушная, полость, окруженная клетками мякоти листа. Клетки эпидермы, примыкающие к замыкающим, получили название побочных, или околоустьичных . Они участвуют в движении замыкающих клеток. Замыкающие и побочные клетки образуют устьичный аппарат .
Число и распределение устьиц на листе или побеге варьируют в зависимости от вида растений и условий жизни. Число их обычно колеблется от нескольких десятков до нескольких сотен на 1 кв.мм поверхности.
Механизм движения замыкающих клеток весьма сложен и неодинаков у разных видов. У большинства растений при недостаточном водоснабжении в ночные часы, а иногда и днем тургор в замыкающих клетках понижается и щель замыкается, снижая тем самым уровень транспирации. С повышением тургора устьица открываются. Считают, что главная роль в этих изменениях принадлежит ионам калия. Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов. Первичный крахмал хлоропластов, превращаясь в сахар, повышает концентрацию клеточного сока. Это способствует притоку воды из соседних клеток и переходу замыкающих клеток в упругое состояние.
Общая площадь устьичных отверстий составляет лишь 1-2% площади листа. Несмотря на это, транспирация при открытых устьичных щелях достигает 50-70% испарения, равного по площади открытой водной поверхности.